
Birds are a subset of dinosaurs (See: Honey, I Shrunk the Dinosaurs. Prior to the extinction of all of the other dinosaurs, resulting from a cosmic impact event about 65 million years ago, the ancestors of modern birds were different from modern birds in many ways, but also similar in many ways. Many of those similarities were features that were actually found among many different dinosaurs, and used in ways that modern birds probably don’t use them (see: Birdish adaptations in dinosaurs: Aerosteon riocoloradensis. Giant tyrannosaur like dinosaurs had systems for efficient breathing that modern birds use to facilitate the aerobically demanding task of flight, but those dinosaurs did not fly (see: Aerosteon riocoloradensis: A Very Cool Dinosaur from Argentina. Many dinosaurs had feathers covering or adorning their bodies, but again, modern birds use feathers in ways that overlap with but are different from how those early non-bird dinosaurs used them (see: On The Threshold Of Flight. Similar comparisons can be made with other bird features such as how their hips work.
One of the few features that modern birds have but that was not found in other dinosaurs, or for that matter, early birds, is the beak. The beak is considered by evolutionary biologists to be a “key trait.” The feature distinguishes birds from other species; All birds have them, no other species do. Beaks probably facilitated many other bird adaptations, mainly but not exclusively dietary; Modern birdness comes in part from being beaked. Beaks vary across different kinds of birds, and this great diversity in beaks is part of the great diversity of the 10,000+ species of birds that exist today; Beaks thus facilitated the diversification of birds. Within species, or closely related groups of species, beaks vary by small amounts that result in important adaptive fine tuning, as we see in the Grand’s studies of the Galapagos Finches; Beak related behavior is at the cutting edge of survival for many bird species.
See: Four Wings Good Two Wings Better?
So how did beaks evolve? Recent research provides some important information necessary to begin to address this question, but this and other parallel research, on the other end of birds (the tail) also serve to tell us something very important: when it comes to understanding how evolution actually happened (of birds or anything else) we are standing at the very start of a long and uncertain journey. In other words, we are nearly clueless.
Years ago a famous writer turned the idea of finding ancient, possibly dinosaur, DNA and turning it into a theme park with dinosaurs grown in modern times. Around the same time, dinosaur experts, such as the famous Jack Horner, postulated that all of the DNA needed to make a modern day dinosaur probably resided, in adjusted, re-used, or silenced form, in living birds. If only you could make fine adjustments to the expression of existing DNA you could hatch a dinosaur, starting with, for example, a turkey.
See: Are Birds Really Dinosaurs?
That was a long time ago and we’ve made nearly zero progress in this area. Meanwhile, GMO technology, mainly for plants, has been developed to produce amazing vegetables that have amazing properties and solve major problems. Except, really, that hasn’t happened either. GMOs are interesting, but like the Platypus (according to Phineas and Ferb), they don’t to much. So this new research is very interesting, and we applaud the scientists for their work. But this work also serves to remind us that the processes of genetic change that results eventually in new species, and the occasional adaptive radiation of myriad species (like the birds), is about as well understood by science as the ecology of a rain forest is understood by a grub that greedily eats a tiny portion of one of the trees in that forest. Well, maybe a little better than that but not much.
See: What Happened With Archaeopteryx?
Bhart-Anjan S. Bhullar, Zachary S. Morris, Elizabeth M. Sefton, Atalay,Tok, Masayoshi Tokita, Bumjin Namkoong, Jasmin Camacho, David A. Burnham and Arhat Abzhanov’s paper, “A molecular mechanism for the origin of a key evolutionary innovation, the bird beak and palate, revealed by an integrative approach to major transitions in vertebrate history,” was just published in the journal Evolution.
These researchers carried out two different major efforts that in combination advance our understanding of the evolution of the bird beak. First, they looked very closely at the development of the beak or snout of several species, including birds and a number of more distantly related forms like crocodilians, and at the same time, looked at dinosaur bones. This formed the basic comparative anatomy and ontogeny that allowed them to specify which bird bits corresponded to which non-bird bits. This is an interesting topic whenever you run into it. I enjoy, for example, telling students that part of the mammalian pituitary gland was originally part of the top of the mouth, and another part originally part of the bottom of the brain, but during development they got together and formed this new feature. (The students seem rarely impressed or interested, but I enjoy telling them anyway.) Along with this ontogenetic reconstruction, the researchers also identified two genetic pathways that are associated with this developmental process.
The second thing they did was to interfere with the genetic pathways in some chickens, which produced a chicken with beaky bits that were more like snouty bits found in, for example, a Alligator. Like this:
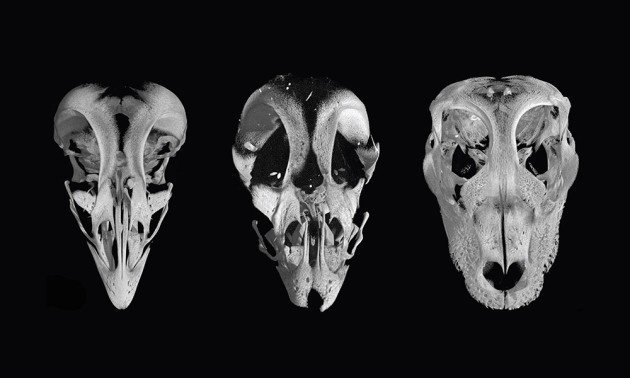
Apparently it is ethically questionable to create a genetically modified chicken and then hatch it, so approval for that research wasn’t even sought at this stage. The chickens were done in before hatching. Most likely, the researchers have indicated, the hatched birds would be viable and not too different from regular chickens, possibly less different than some of the odd breeds generated by more traditional methods.
Here’s the thing. The genetic pathway found in Alligators and birds and everything within the containing family tree of Alligators and birds was messed with, and the result was the loss of bird-osity and thus reversion to alligatorness. However, both birds and Alligators actually use this pathway. Alligator are not birds lacking this pathway. The difference between birds and non birds, with respect to the beak, is almost certainly not in the presence of absence of this pathway, but in other important details, such as signaling or control genes, that determine exactly how the genetic pathway is used. Figuring that out would be a next step in this research, and will likely prove difficult. There have been a number of research projects (including the aforementioned project of figuring out how birds came to have reduced tails) that give a clue as to which genetic and developmental systems are altered in evolutionary time to distinguish among species, but that clue tends to lead to more questions than it answers. From a pragmatic point of view, this means that if you are young and in school and want to eventually get a PhD and study this evolutionary problem, you can probably do that; This won’t be figured out between now and the time you get your first grant!
The paper is available on line, but for your convenience, here is the abstract:
The avian beak is a key evolutionary innovation whose flexibility has permitted birds to diversify into a range of disparate ecological niches. We approached the problem of the mechanism behind this innovation using an approach bridging paleontology, comparative anatomy, and experimental developmental biology. First we used fossil and extant data to show the beak is distinctive in consisting of fused premaxillae that are geometrically distinct from those of ancestral archosaurs. To elucidate underlying developmental mechanisms, we examined candidate gene expression domains in the embryonic face: the earlier frontonasal ectodermal zone (FEZ) and the later midfacial Wnt-responsive region, in birds and several reptiles. This permitted the identification of an autapomorphic median gene expression region in Aves. In order to test the mechanism, we used inhibitors of both pathways to replicate in chicken the ancestral amniote expression. Altering the FEZ altered later Wnt responsiveness to the ancestral pattern. Skeletal phenotypes from both types of experiments had premaxillae that clustered geometrically with ancestral fossil forms instead of beaked birds. The palatal region was also altered to a more ancestral phenotype. This is consistent with the fossil record and with the tight functional association of avian premaxillae and palate in forming a kinetic beak.
________________________
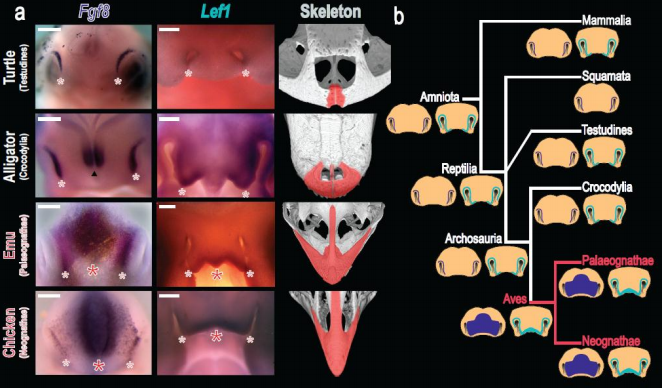
Caption for image at top of post: Figure 2. Expression of facial patterning genes across amniotes. a, Expression of Fgf8 and Lef1 and adult skeletal phenotypes in reptiles, showing ancestral paired gene expression preceding small, paired premaxillae in turtles and alligators and median zone of expression preceding elongate, fused premaxillae in birds. Skeletal images from UTCT/digimorph.org b, Amniote phylogeny with data on facial patterning gene expression and inferred ancestral states shown. Dark blue: Fgf8. Light blue: Lef1. The median expression zone is an autapomorphy of birds correlated with the presence of the avian rostrum. Scale bars 500 ?m.












Leave a Comment